
It is well known that the Risø mutant 1508 (Ingversen et. al., 1973) with the gene lys-3a (Jensen and Doll, 1979) has a content in lysine about 44% over its parental variety Bomi and so a lower yield.
In 1978, initiated a breeding program for high-lysine barley with 24 European commercial varieties, 18 winter barleys and 6 spring barley varieties, introducing the gene lys-3a of Risø 1508. Along the work, we selected the high- lysine progenies by the ninhydrin colour test (Mertz et. al., 1974) after each self-generation. At the same time, in order to check the efficiency of this colorimetric method, we analyzed the same seeds for protein and lysine content. Protein content was determined by gas chromatography using a model ANA 1400 nitrogen analyzer; lysine content was determined by ion-exchange chromatography using a Carlo Erba model 2A27 amino acid analyzer.
In this paper, we report lysine and protein contents of five BC2F2 in comparison to their parent varieties (see Table 1). When we determined the basic amino acids content by ion- exchange chromatography, we found a great variation in the increase of lysine content in four from the five BC2F2 progenies in relation to its parental varieties and, we noticed, also an important variation of ammonia content. Because of this, we calculated the relationship lysine/ammonia in order to compare our results with the Hans Doll's finding (1973). All data of parents and selected lines were compared by t-test. Non-significance and significance at P<0,05, P<0,01 and P<0,001 are indicated with N.S.9 *1 **, and ***, respectively. Also, we present the amino acid composition of two BC2F2 and 100 kernel weight of four BC2F2 (see Tables II and III).

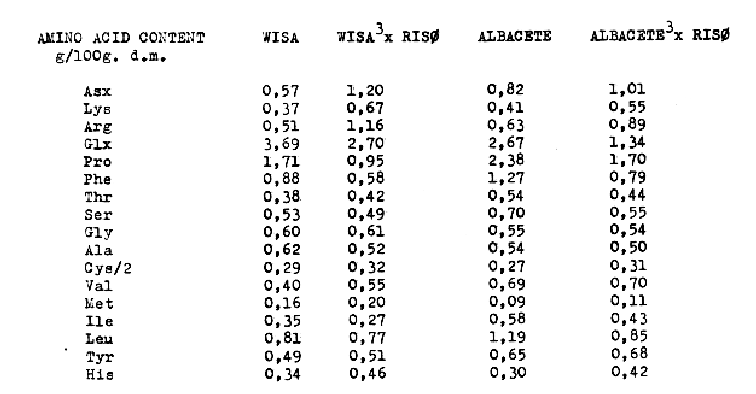

From the results presented in this report we conclude:
References:
Doll, H. 1973. Inheritance of the high-lyBine character of a barley mutant. HereditaB 74:293-294.
Doll, H., Andersen, A. J., Koie, B. and Ingversen, J. 1973. High lysine mutants in barley. Barley Genetics Newsletter 3:12-13.
Ingversen, J., Koie, B. and Doll, H. 1973. Induced seed protein mutant of barley. Experientia 29:1151-1152.
Jensen, J. and Doll, H. 1979. Gene symbols for barley high- lysine mutants. Barley Genetics Newsletter 9:33-37.
Kreft, I., Javornik, B. and Kajfez-Bogataj, L. 1985. Ripening and yield in barley with high-lysine gene lys- 3a. Barley Genetics Newsletter 15:64-67.
Muench, S. R., Lejeune, A. J., Nilan, R. A. and Kleinhofs, A. 1975. Evidence for two independent high lysine genes in barley. Barley Genetics Newsletter 5:31.
Munck, L. 1972a. High lysine barley- a sumary of the present research development in Sweden. Barley Genetics Newsletter 2:54-59.
Tallberg, A. 1973. Ultrastructure and protein composition in high-lysine barley mutants. Hereditas 75:195-200.
Tallberg, A. 1980. Comparison between screening methods for lysine with the use of a barley material with a varying amino acid composition. Acta Agriculturae Scandinavica 30:26-32.
Tallberg, A. 1982. Characterization of high-lysine barley genotypes. Hereditas 96:229-245.
Welch, R. W. 1978. protein barley genotypes. Cereal Research Communications, vol. 6, N. 4:393-398. Evaluation of high-lysine and high-