
Abstract. Two short-awned mutants, 'mt1' and 'mt2' Of varieties 'HBL 175' and 'HBL 176' respectively were allelic and controlled by the same genes since their F1 population gave short-awned plants and there was no segregation in F2 and F3 generations. Similarly, mutants for awnless and blue aleurone were allelic and controlled by single genes.
Introduction. Awnlength varies considerably between spikelets and spikes of single plants and between barley varieties. Monofactorial recessive inheritance of short- awned genes over long-awned had been reported by Helgason (1963), Gaul and Hesemann (1966) and Takahashi and Hayashi (1982). Eslick and Hockett (1967) reported that the genes controlling awn length in seven mutants were allelic.
Blue aleurone colour in barley is due to anthocyanin pigment in some or all aleurone cells. It is best seen in well filled grains magnified to show individual aleurone cells. Colour expression varies with environment and is genetically modified (Finch and Simpson, 1978).
The objectives of the present study were: (a) to ascertain the number of genes controlling expression of short-awned, awnless and blue aleurone mutant phenotypes and (b) to ascertain the allelic relationships of these genes.
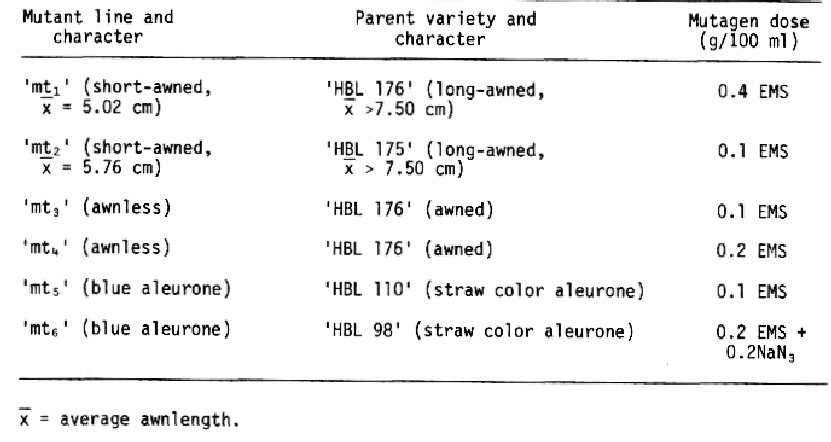
Materials and Methods. The origin of the mutant lines is presented in Table 1. Information regarding allelism of genes was obtained from F1 phenotypes and F2 segregation patterns. The number of genes was determined by comparing observed with expected F2 ratio. Breeding behaviour of the F2 was confirmed in the F3.
Results and Discussion. The cross of short-awned mutants (average awnlength 0.50 cm), 'mt1' × 'mt2' gave short-awned F1 plants (average awnlength = 4.98 cm), suggesting that the genes controlling the two mutants were allelic. The F2 population consisted of 35 short-awned plants (average awnlength 4.22 cm), revealing the absence of segregation between the genes controlling the character. Similarly, crosses between awnless mutants 'mt3' and 'mt44' and blue aleurone mutants 'mt5' and 'mt6' resulted in F2 and F3 progeny with no segregation, indicating that genes for these characters were allelic and controlled by single genes (Table 2). Earlier workers, Eslick and Hockett (1967) and Tsuchiya (1974a) also reported the presence of allelic genes for short-awned mutants. Kushnak (1974) also reported allelic nature for the genes Blx with Bly of blue aleurone whereas, Finch and Simpson (1978) and Genozalez Moreno (1981) found presence of complementary dominant genes and dominant epistatis respectively for the expression of blue aleurone.

References
Eslsick, R. F. and E. A. Hockett. 1967. Allelism for awn length Lk2 in barley (Hordeum species) Crop Sci. 7:266-267.
Finch, R. A. and E. Simpson. 1978. New colours and complementary colour genes in barley. Pflanzenz. 81:40-53.
Gaul, H. and C. W. Hesemann. 1966. Importance of macro- mutation for plant breeding. I. Examples of awn length in spring barley mutants in a different genetic back- ground. Z. Pflanzenz. 55:225-237.
Genozalez Moreno, A. 1981. The inheritance of "blue aleurone endosperm" character in barley. Anales del Institute Nacional de Investigaciones Agraries Agricola 15:33-36.
Helegason, S. B. 1963. Inheritance of glume characters in some barley crosses. Proc. lst Int. Barley Genet. Symp. Wageningen, 26-31 August, 1963-64. 181-185.
Kushnak, G. D. 1974. Utilizing linkages of genetic male sterile and aleurone colour genes in hybrid barley. (Hordeum vulgare L.) systems. Dist. Abst. Int. B 35:148413.
Tsuchiya, T. 1974a. Allelic relationships of genes for short-awned mutants in barley. Barley Genet. Newsletter. 4:80-81.