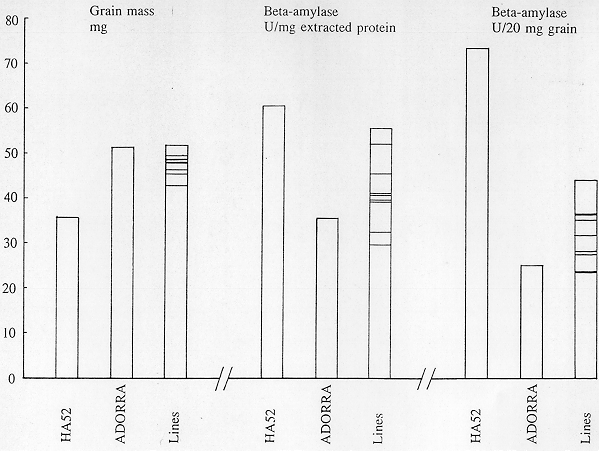
Figure 1. Grain mass and beta-amylase on extracted protein or grain bases in HA52, Adorra and nine lines of their intercross.
Introduction
Previously, we have shown that genetic resources of high total beta-amylase activity exist in some wild Hordeum spontaneum barley strains (Ahokas & Naskali 1990a,b). Genes responsible for this activity could be recovered in domesti- cated phenotypes from backcross (BC-F6) descendents with cv Adorra (H. vulgare) as determined by activity measurements and Southern blotting (Ahokas & Erkkilð 1992). However, due to selection for domesticated phenotype, the genes derived from H. spontaneum tend to be lost. Beta-amylase activity in H. spontaneum shows a strong negative correlation with the studied moisture parameters of the original habitats in Israel and Jordan: with annual rainfall, r=-.860 (P<.001) and annual number of rain days, r=-.912 (P<.001) (Ahokas & Naskali 1990b). This suggests that high beta-amylase content in protein is best adapted to arid habitats within the species distribution. Chalmers et al (1992) likewise found rain day numbers and January temperatures to explain over 78 percent of the variation in a particular beta-amylase isozyme occurring in arid habitats in Isreal.
We have repeatedly transferred genes responsible for high beta-amylase activity from high-activity strains of H. spontaneum, with single cross or backcross, with cv Camir (ant13-400) or cv Adorra, to vulgare-like segregants (results not shown).
The variably shrunken, semifertile, high-lysine mutant HA52 (Ahokas 1977) selected from a Finnish landrace, and F5 lines selected for plump grains and perfect fertility of the cross HA52/Adorra were analyzed. This interest arose from the earlier reports that the shrunken/high-lysine cv Hiproly has a high level of free beta-amylase (Allison & Swanston 1974; Hejgaard et al 1979). HA52 was found to carry high total beta-amylase activity, which is principally not asso- ciated with shrunkeness or semifertility. Stressful conditi- ons may induce leaf pigments defect in HA52, whose shrunk-eness and semifertility are also environmentally sensitive.
Material and methods
The material were field grown, fertilized with 500 kg/ha (N-P-K, 16-7-13.3) in single row plots side-by-side, planted and harvested by hands in SE Finland in the 1993 season. Husked and surface sterilized grains were germinated under axenic conditions in darkness for five days. A sample of fifteen germinating grains were extracted and analyzed aseptically as described before (Ahokas & Naskali 1990a, Ahokas & Erkkilð 1992): for total beta-amylase using the Betamyl method (Megazyme) and for protein in the extracts using the bichinconinic acid method (Sigma). After five days' germination at 15C, the activity is expected to show the total activity because of the natural release of the bound form (Grime & Briggs 1995).
Results
The variably shrunken, semifertile (134 plants) vs normal (79 plants) segregation in F2 of the cross HA52/Adorra does not fit the 3:1 ratio (X2=16.603; P<.001), but fits the 11:5 (X2=3.380; P>.05). This may mean that two complementary genes are involved. Likewise, the F1 spikes of HA52/Adorra grown in the field also segregated 12 shrunken and 132 plump grains plus 5 sterile florets. The shrunken vs plump ratio fits a 1:15 ratio (P>.30).
After selection for better stem and grain type, nine lines were retained and subjected, as F(6) seeds, to the beta-amylase determination with HA52 and cv Adorra grown together. The total beta-amylase activity in HA52 were, on the extracted protein basis, 1.7 times and, on the mass basis, 2.9 times that in cv Adorra (Figure 1). In two lines of the cross HA52/Adorra, the level of activity on the extracted protein basis in HA52 was almost reached in plump-grained fertile segregants the majority being intermediary, while on the grain basis, the activity in cv Adorra could be doubled in the intercross lines (Figure 1). The shrunken-semifertile HA52 background probably still increases the beta-amylase level. These lines showed no (negative) correlation between grain mass and beta-amylase activity (r=.259, on grain basis; r=.519, on extracted protein basis; not significant). This also suggests that high beta-amylase activity is determined by another locus than shrunkeness. Some heterozygotes controlling beta-amylase may have still occurred among the lines.
Discussion
HA52 was selected from a landrace by the first author in 1974. This landrace possibly originates from the Jððski municipality, in the former SE Finland, and was brought along by Karelian refugees in 1944. Finnish landraces, practically lost now, were never systematically studied for beta-amylase. However, a spontaneous high beta-amylase cv Olli (Allison & Swanston 1974) was directly selected from a landrace from the Kurikka municipality in W Finland. Cv Olli was released in 1927 without any malting trials (Sauli 1927, 1930) but was widely grown for malt in various regions of Canada before 1984 (Wolfe 1988). Cv Olli, with about 50 percent of its beta-amylase activity in bound form belongs to the low free form class (Bendelow 1964), or intermediary class (Allison & Swanston 1974). Also cv Pirkka (a4459), a high beta-amylase barley (Simberg 1950; Allison & Swanston 1974) released in 1952, has two of its four crossed parents from Finnish landraces (Simberg 1950; Kivi 1969).
It may be argued that something in the past agriculture in Finland has been selective for high beta-amylase. Barley cultivation dates back to more than 3000 years in Finland (Vuorela & Lempiðinen 1988), barley being the most important cereal until the eighteenth century (Grotenfelt 1922). In the past, barley was usually cut quite early, at yellow maturity, and was dried before threshing, often on various kinds of wooden hurdles (Grotenfelt 1922). He reasoned that the premature harvesting was to avoid shattering. However, fear of night frosts, earlier planting of a successive crop, winter rye or shortage in previous cereal stores may also have driven people to harvest too early. It is also possible that premature harvesting improved the end-use quality in food-making.
The premature cutting subjected the living barley stems to water deficit. The occurrence of a high level of beta-amylase, a stored protein of grain, is highly correlated with drought parameters in the habitats of H. spontaneum (Ahokas & Naskali 1990b). The premature harvesting and drying in past cultivation, possibly for hundreds of generations in Finland, may have promoted the frequency of high beta-amylase genotypes, especially in the latest maturing plants of the populations. Likewise, the tradition- al premature barley harvesting, mainly when the plants were still green, was linked by Bothmer et al (1990) to the occurrence of the brittle rachis in mature spikes in Tibet.
References
Ahokas, H. 1977: Increase in protein content by partial fertility. - Barley Genet. Newslett. 7: 6-8.
Ahokas, H. & Erkkilð, M. J. 1992: Barley beta-amylase and beta-glucanase activities at germination in vulgare-type lines from backcrosses of wild, spontaneum strains with cv. Adorra. - Agric. Sci. Finl. 1: 339-350.
Ahokas, H. & Naskali, L. 1990a: Variation of alpha-amylase, beta-amylase, beta-glucanase, pullulanase, proteinase and chitinase activity in germinated samples of the wild progenitor of barley. - J. Inst. Brew. 96: 27-31.
Ahokas, H. & Naskali, L. 1990b: Geographic variation of alpha-amylase, beta-amylase, beta-glucanase, pullulanase and chitinase activity in germinating Hordeum spontaneum barley from Israel and Jordan. - Genetica 82: 73-78.
Allison, M. J. & Swanston, J. S. 1974: Relationships between beta-amylase polymorphisms in developing, mature and ger- minating grains of barley. - J. Inst. Brew. 80: 285-291.
Bendelow, V. M. 1964: Inheritance of free beta-amylase in barley. - Canad. J. Plant Sci. 44: 550-554.
Bothmer, R. von, Yen, C. & Yang, J.-L. 1990: Does wild, six- rowed barley, Hordeum agriocrithon, really exit? - FAO/IBPGR Plant Genet. Resour. Newslett. 77: 17-19.
Chalmers, K. J., Waugh, R., Watters, J., Forster, B. P.,
Nevo, E., Abbott, R. J. & Powell, W. 1992: Grain isozyme and ribosomal DNA variability in Hordeum spontaneum po- pulations from Israel. - Theor. Appl. Genet. 84: 313-322.
Grime, K. H. & Briggs, D. E. 1995: Release and activation of barley beta-amylase. - J. Inst. Brew. 101: 337-343.
Grotenfelt, G. 1922: Suomalainen peltokasviviljelys. II. - Otava, Helsinki, 517 pp.
Hejgaard, J., K½ie, B., Karlsson, K.-E. & Tallberg, A. 1979: Beta-amylase activity - A simple screening test in Hiproly barley breeding. - Hereditas 90: 145-147.
Kivi, E. I. 1969: Main features of agricultural plant breeding in Finland. - Peat Plant News 1(4): 45-53.
Sauli, J. O. 1927: Suomen maatiaisohrat ja niiden jalostusarvo. (German summary: Die finnischen Landgersten und ihr zchterischer Wert.) - Suomen Maataloustieteel- lisen Seuran Julkaisuja 16: 1-139.
Sauli, J. O. 1930: Selostus Tammiston uusimmista kauppaan lasketuista jalosteista. - Siemenjulkaisu 1930: 180-183.
Simberg, N. H. 1950: Unders—kning av maltkorn. - Diploma Thesis, Univ. of Technol., Dept. of Chem. Engin., Helsinki, 118 pp.
Vuorela, I. & Lempiðinen, T. 1988: Archaeobotany of the site of the oldest cereal grain find in Finland. - Ann. Bot. Fenn. 25: 33-45.
Wolfe, R. I. 1988: Areas devoted to barley varieties in western Canada and the Peace River region 1973-1987. - Barley Newslett. 31: 216-218.
Figure 1. Grain mass and beta-amylase on extracted protein or grain bases in
HA52, Adorra and nine lines of their intercross.